 |  |
| 秦田勇二 氏 | 大田ゆかり 氏 |
1 .はじめに、海は深い
驚くことに、海洋の深さ平均はナントナント富士山の高さと同じくらいの数値(3,8 キロメートル)と計算される。海洋の最深部マリアナ海溝に至っては深さ11キロメートルにまで及ぶ。“海は広いーな♪”だけでなく、“海は深いーな”のである。
「深海がどのような地形なのか?深海では何が繰り広げられているのか? 深海にはいったい何がすんでいるのか?」などの興味深い謎を調査するために独立行政法人海洋研究開発機構は深海探査機や有人潜水調査船を保有している。有人潜水調査船「しんかい6500」(図1)はその名の通り、人が乗船したまま6,500mまで潜水することが可能であり、中からマニピュレーターを操り(図2)、研究者が目視して興味深い環境の土壌・生物などを直接サンプリングすることができる。探査機・調査船を用いた調査の結果、深海には想定していた以上の生物量が存在していることが明らかとなり、深海に生息する微生物の新規性や多様性も大変高いことが判明してきた。
 |  |
| 図1. 有人潜水調査船「しんかい6500」 | 図2. マニピュレータによる深海底泥サンプルの採取 |
2. カラギーナン分解酵素
2-1. 基質の違いを厳密に区別
日本では古くから食品の増粘剤やゲル化剤として使用される多糖にカラギーナンがある。カラギーナンは、ツノマタ、キリンソウなどの紅藻類の細胞壁から抽出される。カラギーナンの特徴は硫酸基を持っている点にあり、その硫酸基パターンに基づき、カッパ、イオタ、ラムダ型の3タイプに大別される(図3)。それぞれの物理的性質が大きく異なっているため、用途に応じて使い分けがなされている。カラギーナンは、原料藻の品種や採取地、生育段階によって、その置換基パターンが多種多様に変化する。このため、いわゆるカラギーナンという名称で市場に流通している物は、実際には非常にヘテロな高分子構造を有しており、精密な構造解析はかなり困難である。そこで我々は酵素の“基質特異性”を生かした分析手法の開発に取り組んだ。
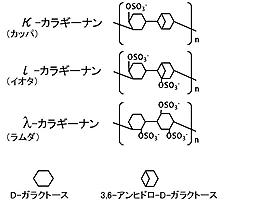
カラギーナン分析手法の確立には、主要な3タイプのカラギーナンそれぞれに対して各構造を厳密に区別して認識する機能、すなわち高度の基質特異性を持つカラギーナン分解酵素3種を利用できることが鍵であった。その3種の酵素を“新規性と多様性の高い”深海微生物から取得することに成功した。(参考文献1、2)。驚いたことにラムダカラギーナン特異的分解酵素のアミノ酸配列は、データベースに登録されているいずれのタンパク質にも相同性を示さず、新規酵素種としてInternational union of biochemistry and biology(IUBMB)に認められ、新しいECナンバー3.2.1.126が付与された。世界的にも目新しい大きな発見であった。深海微生物の高い新規性と今後の可能性を感じさせられた一例である。
2-2. 3種の酵素で食品素材中のカラギーナン分析
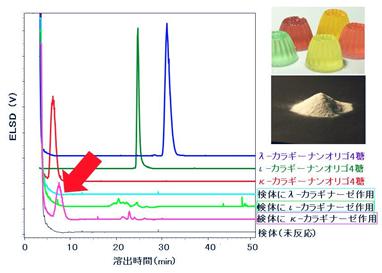
これら3種の酵素を利用した未知カラギーナンの分析法も開発した。この手法は、未知カラギーナンに対し、3種の特異的分解酵素をそれぞれ作用させ、認識部位で限定的に切断して得られるカラギーナンオリゴ糖を分離モードの異なるカラムを使ったHPLC分析でマッピングして、構造・組成を分析するというものである(図4)(参考文献3)。飲食品に含まる未知カラギーナンに対しては、糖質に代表される多量の夾雑物からの妨害を受けるため、有効な特異的分析手段がこれまでなかった。我々の開発した特異的酵素群を用いるこの手法は、飲食品中のカラギーナンの検出・組成分析に極めて有効であった。
3. 寒天分解酵素の利用
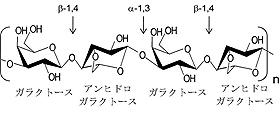
紅藻類の仲間にはテングサやオゴノリなど寒天の主成分であるアガロースを大量に含むものがある。古くから日本ではこれらの紅藻類から寒天(アガロース)を抽出し、食品原料として利用してきた。アガロースは図5に示した通り、D-ガラクトースと3、6-アンヒドロ-L-ガラクトースがα-1, 3、β-1, 4結合で交互につながったヘテロ多糖である。このようにガラクトースが基本骨格となった多糖は陸上の植物からほとんど検出されない。さらに興味深いことに、アガロースをオリゴ糖サイズにまで切断して得られる「寒天オリゴ糖」には、がん抑制効果、免疫機能調節、肝障害改善効果、抗炎症作用、抗リウマチ作用、保湿効果、美白効果(メラニン産生抑制作用)などさまざまな生理的機能があることが報告され、注目されている。著者らはアガロースを目的のサイズのオリゴ糖に切断する手段として、アガロース切断酵素(アガラーゼ)の利用を目指した。アガラーゼは大きく2種類に分類される。アガロースのα-1, 3結合を切断する酵素をα-アガラーゼと呼び、β-1, 4結合を切断する酵素をβ-アガラーゼと呼ぶ。酵素は通常、基質特異性(基質選択性と反応特異性)が大変高いのでα-1, 3とβ-1, 4結合の両方を切断する酵素は存在しない。
深海微生物A33株から検出されたアガラーゼはα-アガラーゼ(α-1, 3結合を切断)であった(参考文献4)。α-アガラーゼの発見例は非常に稀で、本例が世界で2例目にあたる。A33株はその培地中に寒天が含まれている時にだけアガラーゼを生産した。ノリ(海苔)に含まれている多糖はポルフィランと呼ばれ、以前から抗酸化活性、抗アレルギー作用、抗腫瘍活性、免疫賦活作用などを有すると報告されていた。興味深いことに、図6に示す通り、本アガラーゼでポルフィランを切断処理することによってその抗酸化活性が飛躍的に向上した(参考文献5)。
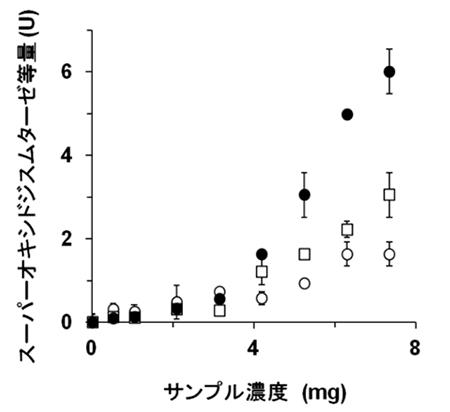
●ポルフィランにαアガラーゼを作用させた
□ポルフィランにβアガラーゼを作用させた
○未処理のポルフィラン
A11株から発見された酵素はβ-アガラーゼであった。本酵素でアガロースを切断すると主産物としてネオアガロオリゴ2糖を生成した(参考文献6)。サイズが小さい寒天オリゴ糖であることから、塗布すると皮膚表面からの浸透に有利であると考えられ、深部のメラニン産生細胞に届くことでメラニン産生を抑え、肌の美白効果が期待できる。
A94株から発見されたβ-アガラーゼはとても高い耐熱性を持っていた。例えば60℃の高温中に15分間放置しても活性の低下はほとんど見られなかった(参考文献7)。遺伝子試薬メーカーのニッポンジーン株式会社から本アガラーゼを試薬として開発する提案を頂き、現在、遺伝子研究用試薬として販売されている(図7)。本アガラーゼを用いると高分子(160 kbpまで確認済み)のDNAが効率よく回収・調製できることから、さまざまな研究機関で遺伝子の機能解析・配列解析に大きく貢献している。
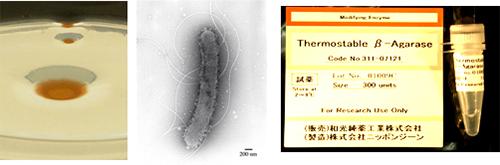
同微生物の電顕写真(中)、耐熱性アガラーゼ製品(右)
4. おわりに、多様な可能性
地球上の炭素の分配を見ても海洋中に含まれる炭素比率は陸上と比較して17倍多いと計算される。生物は海洋から発生し進化を続け、多様性を創り上げてきた。したがって、海洋には数限りない種類の生体触媒(酵素など)が存在している。それらの生体触媒から、人々の生活を豊かにする生体触媒を探し出すことに大きな可能性を感じるのである。
今回紹介させていただいた酵素のほかにトレハロースを生成するための酵素や難溶性物質に糖を結合させて水溶性を向上させることのできる酵素などが得られており、企業との共同開発が進められている。
そのほか、力を入れているのがリグニンを有効利用するための深海微生物の応用研究である。リグニンはセルロースに次ぐ第2の存在量を誇る多大なバイオマスであり、その化学構造から判断すると、さまざまな石油化学製品の代替品に変換することが可能と推定される。深海から回収した腐食の進んだ沈木から多様なリグニン関連芳香族を代謝する微生物を検出した(参考文献8)。さらにリグニン内に多く存在するβ-O-4結合を特異的に切断できる微生物も検出できている。今後はこれらの微生物の生物機能を活用したリグニン有効利用の可能性を追求する。
参考文献
1) Y. Ohta, & Y. Hatada,: J. Biochem., 140, 475(2006)
2) Y. Hatada, et al. : Mar. Biotechnol., 13, 411(2011)
3) 秦田勇二, 大田ゆかり: 財団法人アサヒビール学術振興財団 研究紀要 22, 23(2009)
4) Y. Ohta, et al. : Curr. Microbiol., 50, 212 (2005)
5) Y. Hatada, et al. : J. Agric. Food Chem., 54, 9895(2006)
6) Y. Ohta, et al. : Biotechnol Appl Biochem 41, 183(2005)
7) 秦田勇二等、極限環境生物の産業展開、シーエムシー出版社、190(2012)
8) Y. Ohta, et al. : OJMS, 2, 177(2012)
(はただ ゆうじ)
秦田 勇二(はただ ゆうじ) 氏 プロフィール
1965年広島県生まれ。海洋研究開発機構・海洋生命理工学研究開発センター・深海バイオ応用研究開発グループ・グループリーダー。横浜市立大学大学院生命ナノシステム科学研究科・客員教授。花王株式会社にて衣料用洗剤「アタック」に配合する酵素の開発に従事(1993年~2001年)。海洋研究開発機構(元 海洋科学技術センター)にて深海微生物の応用研究に従事(2002年~現在)。日本農芸化学会会員、極限環境生物学会会員、日本応用糖質科学会会員、マリンバイオテクノロジー学会会員(評議員)。工学博士。
(おおた ゆかり)
大田 ゆかり(おおた ゆかり) 氏 プロフィール
1967年山口県生まれ。海洋研究開発機構・海洋生命理工学研究開発センター・深海バイオ応用研究開発グループ・主任技術研究員。海洋研究開発機構(元 海洋科学技術センター)にて深海微生物の応用研究に従事(2002年~現在)。日本農芸化学会会員、日本応用糖質科学会会員、マリンバイオテクノロジー学会会員、木材学会会員。工学博士。
関連リンク