シワのあるえんどう豆とシワのない丸いえんどう豆をかけ合わせると、丸いえんどう豆ができる。このことをメンデルは、丸い遺伝子がシワの遺伝子より優性であると説明した。では、このような遺伝子の優劣関係はどう決まるのか。“シワの遺伝子(劣性遺伝子)は丸くなる機能を失っているから形として現れない”というのがこれまで考えられてきた仕組みだった。
2016年12月、奈良先端科学技術大学院大学のバイオサイエンス研究科の和田七夕子(わだ ゆうこ)助教と高山誠司(たかやま せいじ)客員教授(現東京大学大学院農学生命科学研究科教授)らの研究チームは、この従来の考えを覆す研究成果を発表した。“劣性遺伝子の発現を抑える物質”の存在を確認したのだ。アブラナ科植物の“ある遺伝子”の傍で後天的につくられる、「Smi2」と呼ばれる低分子RNAの存在である。しかもこのRNAは、塩基配列を微妙に変化させることによって、より複雑な優劣関係を制御するという。遺伝子の優劣関係決定の全く新しい仕組みが見えてきた。

→「Nature Plants 3, 16206 (2016)」。
対立遺伝子に生まれる優劣関係
研究チームが着目したのは、アブラナ科の植物の花粉にある「SP11」という遺伝子だ。この遺伝子には、非常に多くの対立遺伝子が存在する。対立遺伝子とは、対になる性質を決める遺伝子をいう。冒頭のえんどう豆の例では、シワがあるかないかという対になる性質について、①丸い豆をつくる遺伝子と、②シワの豆をつくる遺伝子の2つが対立遺伝子であり、①>②という優劣関係がある。SP11遺伝子には、数十種類もの対立遺伝子があり、機能を失ったものはないのに複雑な優劣関係がある。研究チームはそのうちの4つの対立遺伝子を研究対象に選び、それらの優劣性を解析する中で、優劣関係を決める因子Smi2の特定に成功した。
Smi2が劣性遺伝子の発現をブロックする
実験は非常に複雑だが、ここでは簡略化して紹介しよう。遺伝子はDNAの上にあり、そのすぐそばには、その遺伝子が発現するために転写※1を開始させる転写調節領域がある(下図)。この転写調節領域にさまざまなタンパク質が結合することで、遺伝子の発現が制御(ON/OFFが決定)される。
※1 発現するための転写/発現とは、遺伝子上の情報が翻訳されてタンパク質を合成する一連の過程。また転写とは、その過程の初期の段階で、DNA上の遺伝子配列がRNAに写し取られることをいう。

研究チームは、DNAのSP11遺伝子の領域の近くに、逆位反復配列と呼ばれる回文構造を持つ配列の領域があり、そこから24個の塩基からなるSmi2がつくられることを発見した。またSmi2の塩基配列が、SP11対立遺伝子の種類ごとに少しずつ異なっていて、自己のSP11対立遺伝子に対しては何も作用しないことも分かった(下図)。
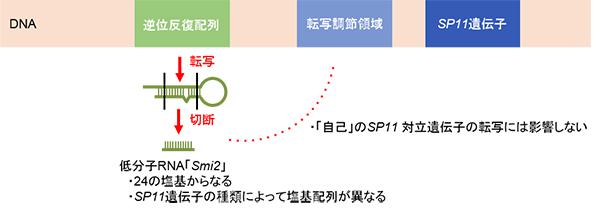
そこで、SP11遺伝子を持つある個体のDNAからSmi2を含む逆位反復配列を取り出し、より劣性のSP11遺伝子を持つ個体に遺伝子導入すると、Smi2は劣性SP11遺伝子の転写調節領域をメチル化※2して制御し、劣性のSP11遺伝子が発現できなくなったのだ(下図上段)。
※2 メチル化/メチル基(-CH3)が付加する反応で、生体内の様々な反応に関わる。
逆に、同じSmi2を、より優性のSP11遺伝子を持つ個体のDNAに入れても、Smi2が転写調節領域をメチル化することはなかった(上図下段)。研究チームは、このSmi2が、劣性の遺伝子の発現を抑制すると結論づけた。
この実験により、冒頭でも紹介したこれまでの遺伝子の優劣関係の概念を覆す仕組みの存在が明らかになった。また、低分子RNAとその標的となる転写調節領域のわずかな塩基配列の違いが、対立遺伝子間に複雑な優劣関係を生むことも分かった。つまり、低分子RNAが進化することを証明したのだ。
一連の実験には多くの要素が盛り込まれ、一つひとつの発見にはさまざまな解析技術が駆使された。ざっと紹介すると、Smi2の発見には次世代シーケンス解析およびゲノムシーケンス、SP11発現調節領域のメチル化はバイサルファイトシーケンス、劣性遺伝子の発現抑制は定量的PCR、といった具合だ。研究チームにうかがうと、特に苦労したのは、Smi2の機能を証明するために、アブラナにSmi2ゲノム領域を導入する実験だったという。研究チームの和田さんは、「Smi2を導入した後も、導入した株と、優劣関係が個々に異なる4種類のSP11を持つ株とを、二回ずつ交配する必要があり、一世代が約半年はかかるので、全ての組み合わせを作出するために数年かかりました」と、多大な時間と労力を費やした実験を振り返る。
「自家不和合性」の謎の解明へ
今回研究チームがSP11遺伝子に着目した背景には、長年取り組んできた自家不和合性の研究があった。自家不和合性とは、植物が種の多様性を保つために、自家受精を防ぐという性質だ。アブラナ科の自家不和合性は、花粉にあるSP11遺伝子が、雌しべにある遺伝子との組み合わせで自己と非自己を認識することで制御される。このSP11に焦点をあてた今回の成果は、自家不和合性の研究にもさらなる可能性を与えたという。
研究チームの高山さんによれば、SP11遺伝子と低分子RNAの関係では、どちらかの変化が引き金となってもう一方に変化をもたらす“共進化”が起きている可能性があり、その過程を明らかにすれば、自家不和合性機構がどのように進化してきたのか分かるかもしれないという。
今回アブラナ科植物で発見されたこのしくみは、他の植物や動物にも存在する可能性がある。“遺伝子の優劣”という生命原理を今後考えていくときの注意点について、高山さんはこう述べる。「優劣性という言葉から、優性の遺伝子が『優(すぐ)れている』と誤解してはなりません。表現型として現れるものを『優性』と呼んでいるにすぎないのです。例えば、緑の革命につながった高収量品種は、ある『劣性遺伝子』を持つ品種です」。
研究が発展すれば、有用な遺伝子を働かせ、有用ではない遺伝子を働かせなくする技術につながるかもしれない。では、何が有用で、何がそうでないのか。例えば、味が良く虫に強い農作物の育種技術はどうだろう。その応用には慎重な議論が必要だろうが、革命的な技術開発の可能性を秘めた成果だと感じた。
(サイエンスライター 丸山 恵)
本文中の解説図3点は、筆者、研究者、編集部で作成